
ЩњЮяЮяРэЫљжмеўзщНвЪОChz1ЪЖБ№ВЂзщзАH2A.ZЕФЗжзгЛњжЦ

ЁЁЁЁдкецКЫЯИАћжаЃЌзщЕААзБфЬхзщзАаЮГЩЬиЪтЕФШОЩЋжЪНсЙЙЃЌетжжгыГЃЙцзщЕААзВЛЭЌЕФШОЩЋжЪНсЙЙЪЧБэЙлвХДЋЕїПиЕФживЊЗНЪНжЎвЛЁЃзщЕААзБфЬхH2A.ZгыГЃЙцзщЕААзH2AОпгаНЯЮЊЯрЫЦЕФвЛМЖађСаЃЌH2A.ZЪЧЖрЯИАћЩњЮяЕФБиашЛљвђЃЌЧвдкЛљвђзЊТМЕїНкЁЂDNAЫ№ЩЫаоИДЁЂЯИАћдіжГЁЂЗжЛЏЕШЙ§ГЬжаЗЂЛгжСЙиживЊЕФзїгУЁЃНќФъРДЃЌЙњПЦДѓВЉЪПЩњЕМЪІЃЌжаЙњПЦбЇдКЩњЮяЮяРэЫљжмеўбаОПзщзлКЯдЫгУНсЙЙЩњЮябЇКЭЩњЮяЛЏбЇЕШЗНЗЈЖдзщЕААзБфЬхЕФЬивьЪЖБ№НјааСЫЯЕЭГбаОПЃЌЯШКѓБЈЕРСЫAnp32eЪЖБ№H2A.ZВЂНЋЦфвЦГ§ШОЩЋжЪЕФНсЙЙКЭЗжзгЛњжЦЃЈCell Research 2014ЃЉЃЈ1ЃЉЃЌвдМАШОЩЋжЪжиЫмИДКЯЮябЧЛљYL1ЪЖБ№H2A.ZВЂАяжњЦфШОЩЋжЪзщзАЕФНсЙЙКЭЗжзгЛњжЦЃЈNat Struct Mol Biol 2016ЃЉЃЈ2ЃЉЁЃ
ЁЁЁЁ5дТ31ШеЃЌжмеўбаОПзщдкPLOS BiologyдгжОдкЯпЗЂБэСЫЬтЮЊStructural insights into histone chaperone Chz1-mediated H2A.Z recognition and histone replacementЕФбаОПТлЮФЃЌНвЪОСЫНЭФИзщЕААзАщТТChz1ЬивьадЪЖБ№зщЕААзБфЬхH2A.ZМАЦфДйНјSWR1ДпЛЏH2A.ZЬцЛЛЕФНсЙЙКЭЗжзгЛњжЦЁЃChz1ЪЧзюдчБЛЗЂЯжЕФФмЙЛЬивьЪЖБ№зщЕААзБфЬхH2A.ZЕФЕААзЗжзгЃЈ3ЃЉЃЌвВЪЧНЭФИЯИАћЬигаЕФH2A.ZзщЕААзАщТТЁЃChz1НсКЯНЭФИЯИАћжагЮРыH2A.Z-H2BЖўОлЬхЃЌВЂФмЙЛДйНјШОЩЋжЪжиЫмИДКЯЮяSRCAP/SWRДпЛЏЕФH2A.ZНЛЛЛЗДгІЃЈ4ЃЉЁЃChz1ШчКЮЬивьадЪЖБ№H2A.ZЃПвдМАШчКЮЭЈЙ§НсКЯгЮРыЕФH2A.Z-H2BЖўОлЬхДйНјSRCAP/SWRДпЛЏЕФH2A.ZНЛЛЛЗДгІЃПетаЉЙиМќЕФПЦбЇЮЪЬтЩаЮДЕУЕННтД№ЁЃ
ЁЁЁЁБОбаОПЪзЯШВтЖЈСЫChz1ЕФCФЉЖЫгыH2A.Z-H2BЖўОлЬхаЮГЩЕФИДКЯЮяЕФОЇЬхНсЙЙЃЌITCЪЕбщБэУїЃЌгыДЫЧАБЈЕРЕФChz1НсЙЙгђ(Chz1-M)ЯрБШЃЌШЋГЄChz1(Chz1-FL)ЖдH2A.ZЕФЪЖБ№ЬивьадЕУЕНДѓЗљЬсИпЃЈЭМ1ЃЉЁЃЦфДЮЃЌбаОПЗЂЯжH2A.ZжаСНИіИпЖШБЃЪиЕФАБЛљЫсВаЛљЃЈGly98КЭAla57ЃЉЖдгкChz1ЬивьЪЖБ№ЕФживЊадЃЌДгЖјНвЪОСЫвЛжжШЋаТЕФH2A.ZЬивьЪЖБ№ЛњжЦЃЈЭМ1ЃЉЃЌетвВЪЧЕквЛДЮдкH2A.ZЕФNЖЫЧјгђЗЂЯжЖдгкзщЕААзЬивьадЪЖБ№ЕФЮЛЕуЁЃзюКѓЃЌSWR1УИЛюВтЖЈЗЂЯжгЮРыЕФH2A.Z-H2BЖўОлЬхвжжЦSWR1ДпЛЏЕФH2A.ZНЛЛЛЃЌЖјChz1ЭЈЙ§НсКЯгЮРыЕФH2A.Z-H2BНтГ§СЫетжжвжжЦЃЌДгЖјДйНјСЫH2A.ZЕФНЛЛЛЗДгІЃЈЭМ2ЃЉЁЃ
 ЭМ1, Chz1-FLЖдH2A.ZЕФЬивьЪЖБ№(a)МАH2A.ZЩЯНщЕМЪЖБ№ЕФЙиМќВаЛљЁё(b)
ЭМ1, Chz1-FLЖдH2A.ZЕФЬивьЪЖБ№(a)МАH2A.ZЩЯНщЕМЪЖБ№ЕФЙиМќВаЛљЁё(b)
ЁЁЁЁБОбаОПВћУїСЫзщЕААзАщТТChz1ЬивьЪЖБ№H2A.ZЕФЗжзгЛњжЦЃЌетжжаТЕФЪЖБ№ФЃЪНгывбБЈЕРЕФЫљгаH2A.ZЪЖБ№ФЃЪНОљВЛЯрЭЌЃЌЭиеЙСЫЮвУЧЖдH2A.ZЬивьЪЖБ№ЛњжЦЕФРэНтЁЃИУбаОПЛЙНвЪОСЫChz1ЭЈЙ§ПижЦгЮРыЕФH2A.Z-H2BЖўОлЬхЕФХЈЖШРДДйНјH2A.ZНЛЛЛЕФзїгУЗНЪНЃЌЩЯЪіЗЂЯжЮЊЩюШыбаОПH2A.ZЕьЖЈСЫЛљДЁЃЈЭМ2ЃЉЁЃ
ЁЁЁЁжмеўПЮЬтзщЙњПЦДѓВЉЪПбаОПЩњЭѕдЦдЦЃЈХрбјЕЅЮЛЃКжаПЦдКЩњЮяЮяРэЫљЃЉЮЊБОЮФЕФЕквЛзїепЃЌжмеўбаОПдБЮЊБОЮФЕФЭЈбЖзїепЁЃИУЯюЙЄзїЕУЕНСЫЛљН№ЮЏЁЂПЦММВПЁЂжаПЦдКBРрЯШЕМзЈЯюЕФОЗбжЇГжЁЃдМКВЛєЦеН№ЫЙДѓбЇЕФCarl WuНЬЪквдМАХІдМжнСЂДѓбЇЪЏЯЊЗжаЃЕФEd LukНЬЪкЮЊИУЯюЙЄзїЬсЙЉСЫАяжњЁЃ
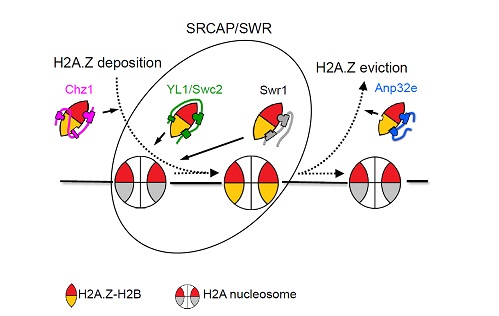 ЭМ2ЃЌВЛЭЌЗжзгЬивьЪЖБ№зщЕААзБфЬхH2A.ZВЂВЮгыШОЩЋжЪзщзАКЭвЦГ§ЕФЛњжЦЮФеТСДНгЃКhttps://journals.plos.org/plosbiology/article?id=10.1371/journal.pbio.3000277
ЭМ2ЃЌВЛЭЌЗжзгЬивьЪЖБ№зщЕААзБфЬхH2A.ZВЂВЮгыШОЩЋжЪзщзАКЭвЦГ§ЕФЛњжЦЮФеТСДНгЃКhttps://journals.plos.org/plosbiology/article?id=10.1371/journal.pbio.3000277
д№Бр ЃКЛЦЧЩ
зюаТЖЏЬЌ
-
2019/09/01ЙужнЕиЛЏЫљдкГЄАзЩНШ§ЭђФъЧАДѓХчЗЂЪТМўЕФГѕДЮРхЖЈКЭОЋШЗЖЈФъШЁЕУбаОПНјеЙ
-
2019/09/01ЙужнЕиЛЏЫљНсКЯдзгСІЯдЮЂОЕКЭФЩУзбЙКлНсКЯдЮЛБэеїВтЖЈвГбвжагаЛњжЪЕФСІбЇаджЪ
-
2019/09/01вХДЋгыЗЂг§ЫљЭѕЙњЖАбаОПзщЗЂЯжВЮгыДѓЖЙдэмеДгАћжЪЕНвКХнзЊдЫЕААз
-
2019/09/01вХДЋгыЗЂг§ЫљИпВЪЯМбаОПзщгІбћдкNature Reviews GeneticsЗЂБэЁАЛљвђзщАаЯђаоЪЮЙЄОпМАЦфдкзїЮяг§жжжаЕФЧАбигІгУЁБзлЪіЮФеТ
-
2019/09/01бЧШШДјЩњЬЌЫљгЁгіСњдКЪПЭХЖгЭъГЩЕкЖўХњжэЬхЯИАћжиРызггеБфЗјееЪЕбщ


